Wide
Distribution of a High-Virulence Borrelia
burgdorferi Clone in Europe and North America
Wei-Gang
Qiu,*
John F. Bruno,† William D. McCaig,* Yun Xu,† Ian Livey,‡ Martin E. Schriefer,§
and Benjamin J. Luft†
*Hunter College of the City University of New York, New York, New York, USA; †Stony
Brook University, Stony Brook, New York, USA; ‡Baxter Innovations GmBH, Orth/Donau,
Austria; and §Centers for Disease Control and Prevention, Fort Collins,
Colorado, USA
Abstract
The A and B clones of Borrelia burgdorferi sensu
stricto, distinguished by outer surface protein C (ospC) gene sequences, are commonly associated with
disseminated Lyme disease. To resolve phylogenetic relationships among isolates,
we sequenced 68 isolates from Europe and North America at 1 chromosomal locus
(16S23S ribosomal RNA spacer) and 3 plasmid loci (ospC,dbpA,
and BBD14). The ospC-A clone
appeared to be highly prevalent on both continents, and isolates of this clone
were uniform in DNA sequences, which suggests a recent trans-oceanic migration.
The genetic homogeneity of ospC-A
isolates was confirmed by sequences at 6 additional chromosomal housekeeping
loci (gap, alr, glpA, xylB, ackA,
and tgt). In contrast, the ospC-B
group consists of genotypes distinct to each continent, indicating geographic
isolation. We conclude that the ospC-A
clone has dispersed rapidly and widely in the recent past. The spread of the ospC-A
clone may have contributed, and likely continues to contribute, to the rise of
Lyme disease incidence.
Multilocus
sequence typing (MLST) is the use of DNA sequences at multiple housekeeping loci
to characterize genetic variations of natural populations of a bacterial
pathogen (1,2).
MLST studies showed that local populations of a bacterial species typically
consist of discrete clusters of multilocus sequence types called "clonal
complexes," rather than a multitude of randomly assorted genotypes (2).
Remaining to be tested are how such factors as natural selection, low
recombination rate, and genetic drift due to geographic structuring contribute
to the formation and maintenance of these clonal complexes in natural bacterial
populations (3,4).
Recently, a multilocus sequence analysis approach was proposed to reconstruct
phylogenetic histories of bacterial clonal complexes by using concatenated
sequences of housekeeping genes when within-loci and between-loci recombinations
are infrequent (5).
Lyme disease is a
multisystem infection, with inflammatory complications that commonly affect the
skin, joints, and central nervous system in humans (6).
Its causative agent, Borrelia burgdorferi,
a spirochete that parasitizes vertebrates, is transmitted by hard-bodied ticks
throughout the temperate zones of the Northern Hemisphere (7).
Although humans are accidental hosts of B.
burgdorferi, Lyme disease is the most common vector-borne disease in
the United States with >20,000 annual reported cases, 93% of which occurred
in 10 northeastern, mid-Atlantic, and north-central states (8).
Small mammals such as white-footed mice (Peromyscus
leucopus) and eastern chipmunks (Tamias
striatus) serve as the main reservoirs of B.
burgdorferi (9,10).
In Europe, B. burgdorferi is
transmitted by Ixodes ricinus
ticks (11)
and is carried by a large variety of hosts, including birds and small- to
medium-sized mammals (12).
B.
burgdorferi sensu stricto is the primary pathogen of Lyme disease in the United
States and is the only pathogenic genospecies that causes Lyme disease in both
North America and Europe. More than 12 distinct outer surface protein C (ospC)
major sequence types coexist in local B.
burgdorferi sensu stricto populations in the northeastern United
States (1315).
Sequence variability at ospC is
the highest among known genomic loci and is strongly linked to variations at
other genome-wide loci, with occasional recombinant genotypes caused by plasmid
exchanges (1619).
B.
burgdorferi sensu stricto intraspecific clonal complexes may differ in their
host specificity and degree of human pathogenecity. Different clonal complexes
may prefer different host species (9).
A restriction fragment length polymorphism type of intergenic spacer (IGS)
sequence (corresponding to the ospC-A
and -B groups) is associated with hematogenous dissemination in patients with
early stage Lyme disease (20,21).
Four ospC clonal complexes (A,
B, I, and K groups) were found to be more likely than others to cause
disseminated Lyme disease (22).
Also, an association of ospC
clonal types with invasive disease in humans has been found in other pathogenic
genospecies such as B. afzelii
and B. garinii (23,24).
However, additional ospC clonal
types have been isolated in patients with invasive disease (14).
Previous
molecular assays found a close relationship and overlapping genotypes between
the European and North American populations (2527).
These authors found greater genetic diversity among American strains than
European strains and proposed a North American origin for this genospecies.
Although these studies provided the first evidence for recent intercontinental
migrations, they left the phylogenetic relationships among clonal complexes
unresolved because of the use of either anonymous genome-wide markers (e.g.,
arbitrarily primed PCR), genes with a high recombination rate (e.g., ospC),
or sequences at a single locus. A phylogeographic approach with multiple
molecular markers provides a more robust inference on population history (28).
Here we obtained a well-resolved phylogeny of B. burgdorferi sensu stricto clonal complexes by using
multilocus sequence typing at housekeeping loci as well as loci under adaptive
evolution. We found evidence of genetic endemism, recent migration events, and
recombinant genomic types. In fact, the highly pathogenic ospC-A
clone seems to have spread rapidly in recent years to infect a broad range of
host species in 2 continents.
Materials
and Methods
B.
burgdorferi Isolates and DNA Isolation
The B.
burgdorferi sensu stricto isolates were obtained from clinical and
tick specimens and cultures from animals in the United States and Europe and
maintained as frozen stocks at 70°C (Table 1). For
in vitro propagation, a small amount of frozen culture was scraped from the
surface of each sample with a sterile inoculating loop and injected into
complete Barbour-Stoenner-Kelly II medium (Sigma-Aldrich Corp., St. Louis, MO,
USA). Spirochetes were then cultivated at 34°C. All cultures used in this study
had undergone a maximum of 2 in vitro passages after recovery from frozen stock.
For isolation of genomic DNA, 10 mL of low-passage log-phase bacteria was
harvested by centrifugation at 10,000 rpm for 30 min at 4°C. The bacterial
pellet was washed twice with Tris-Cl buffer (10 mmol/L Tris [pH 7.5], 100 mmol/L
NaCl), and resuspended in 430 μL TES (10 mmol/L Tris [pH 7.5], 100 mmol/L
NaCl, 10 mmol/L EDTA). Subsequently, 10 μL of freshly prepared lysozyme (50
mg/mL), 50 μL Sarkosyl (10%), and 10 μL proteinase K (10 mg/mL) were
then added, and the mixture was incubated at 50°C overnight before RNase
treatment. After incubation, DNA was extracted with phenol/chloroform and
chloroform, precipitated with ethanol, and finally resuspended in TE buffer (1
mmol/L Tris [pH7.5], 1 mmol/L EDTA).
Genomic
Markers, PCR Amplifications, and DNA Sequencing
PCR
amplifications were attempted at 4 genomic loci for all isolates and at 6
chromosomal housekeeping loci for a genetically representative subset of
isolates (Table 2). The IGS locus was chosen for its
phylogenetically informative polymorphisms (16,20).
The IGS locus and 6 housekeeping genes (gap,
alr, glpA, xylB, ackA,tgt)
were approximately evenly distributed on the main chromosome based on the B31
genome (29).
The 3 plasmidborne loci were selected for their high sequence variability and
for the absence of close paralogs based on a genome comparison (17,19).
IGS sequences were amplified by using a nested PCR procedure (30).
Because of high sequence variability, dbpA
sequences were amplified by using 2 alternative forward primers. PCR
amplification was performed in 50 μL containing 200 mmol/L of each dNTP,
2.0 mmol/L MgSO4, 2.5 U of Platinum Taq DNA polymerase High Fidelity
(Invitrogen, Carlsbad, CA, USA), 0.5 μmol/L of each primer, and 100 ng of
genomic DNA template. Following denaturation at 94°C for 1 min, samples
underwent 30 cycles of denaturation at 94°C for 30 s, annealing at 55°C for
30s, initial extension at 68°C for 1.5 min, and a final extension step at 68°C
for 10 min. PCR products were purified by GFX chromatography (Amersham Pharmacia
Biotech, Inc., Piscataway, NJ, USA), resolved by agarose gel electrophoresis,
and visualized by ethidium bromide staining. Purified amplicons were sequenced
by using standard dideoxy terminator chemistry as outlined below with the
forward and reverse PCR primers. Absence of specific PCR products, indicating
potential absence of particular genetic loci or plasmids, was confirmed by
follow-up amplifications of the flanking DNA segments.
Automated DNA
sequencing of both strands of each fragment was performed by the Stony Brook
University Core DNA Sequencing Facility (Stony Brook, NY, USA) by using the
dye-terminator method with the same oligonucleotide primers used for PCR
amplification or, where required, appropriate internal primers. Sequences were
inspected and assembled with the aid of the Sequencher program (Gene Codes,
Inc., Ann Arbor, MI, USA). DNA sequences were analyzed by using the BLASTN
program through GenBank at the National Center for Biotechnology Information (www.ncbi.nlm.nih.gov).
Nucleotide and protein sequence alignments were performed with MacVector version
6.5 (MacVector, Inc., Cary, NC, USA). New sequences were deposited to GenBank
under accession nos. EF537321EF537573.
Phylogenetic
Inference and Tests of Population Differentiation
The IGS sequences
were used to resolve intraspecific phylogenetic relationships among B.
burgdorferi isolates (16,20).
Two highly divergent tick isolates from Finland (SV1 and Ri5) were used as
outgroups for rooting the phylogenetic tree. IGS sequences were aligned by using
ClustalW (31).
A Bayesian majority-rule consensus tree was estimated by using MrBayes (version
2.1) (32)
as described previously (19).
Sequences at the 3 plasmid-borne proteincoding loci were translated into protein
sequences and aligned in a pairwise fashion with ClustalW (31).
Nucleotide alignments were obtained according to the protein alignments.
Neighbor-joining trees based on pairwise nucleotide sequence distances were
inferred by using PHYLIP (33)
and plotted by using the APE package of the R statistical package (34).
Genetic differentiation among geographic populations was tested by using the
analysis of molecular variance (AMOVA) method implemented in the software
package Arlequin 3.1 (35).
The 6 housekeeping genes were used to infer the overall within- and between-genospecies
phylogeny. Sequences of strains B31 and PBi (B.
garinii) were downloaded from GenBank (29,36).
Sequences of N40, JD1, DN127 (B. bissettii),
and PKo (B. afzelii) were from
draft genomes (S. Casjens, pers. comm.). The 6 alignments were concatenated and
tested for the presence of gene conversion by using GENECONV with the
"within-group fragments only" option (37).
Two approaches, a Bayesian method with codon site-specific evolutionary rates
(using MrBayes) and the other maximum likelihood method with 100 bootstrapped
alignments (using DNAML in PHYLIP) (33),
were used for phylogenetic reconstruction based on concatenated sequences.
Branch supports were measured by the posterior probabilities in the Bayesian
method and the bootstrap values in the maximum likelihood method.
Results and Discussion
AMOVA Tests of
Geographic Differentiation
We sequenced 68
isolates (including 30 from northeastern United States, 6 from the midwestern
United States, and 32 from Europe) at a single chromosomal locus (IGS) and 3
plasmid loci (ospC,dbpA, and
BBD14). Using AMOVA, we evaluated the genetic differentiation among geographic
samples and found significant genetic differentiation between the North American
and European populations at IGS, ospC,
and dbpA, but not BBD14 (Table
3). Among these loci, IGS is the most informative in reflecting the effect
of genetic drift caused by geographic isolation because sequence variations at
IGS are likely to be selectively neutral. In addition, IGS is on the main
chromosome and less likely to be subject to gene conversion. Genetic variations
at 3 plasmid loci are more likely to be influenced by natural selection such as
adaptation to local vector and host species. Also, plasmid genes are more likely
to be transferred so that footprints of geographic isolation might be obscured
by gene flow between populations. Natural selection can both enhance and reduce
geographic differentiation. With adaptation to local habitats, natural selection
acts to enhance the geographic divergence, especially at target loci. On the
other hand, diversifying selection within populations inflates within-population
diversity, which results in lack of differentiation within populations relative
to the within-population polymorphism.
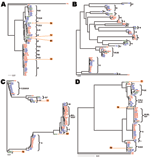
Figure
1. Gene trees showing nucleotide sequence clusters of
68 Borrelia burgdorferi
isolates at 1 chromosomal locus...
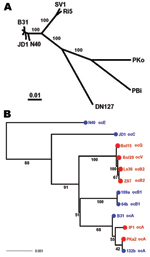
Figure
2
Figure
2. Species phylogeny based on concatenated sequences
at housekeeping loci...
The low level of
geographic differentiation at ospC
showed the divergence-reducing effect of natural selection. Genetic variability
of ospC is as high within
populations as between populations and is caused by diversifying natural
selection (9,13).
In such a case, summary statistics such as AMOVA fixation index (FST)
are misleading because sequence cluster analysis showed that most ospC
alleles have geographically restricted distributions (Figure
1, panel B). The insignificant AMOVA result at BBD14 might be due to a
similar effect of high within-population polymorphisms as a result of
diversifying selection. In contrast, dbpA
showed the divergence-enhancing effect of natural selection. The dbpA
locus showed the highest level of geographic differentiation, owing to a shared
allelic type among B2, L, S, Q, and V clonal groups in Europe (Table
3; Figure 1,
panel C). An adaptive sweep likely has homogenized these divergent European
lineages at dbpA.
In summary, on
the basis of the neutral genetic variations at IGS, we conclude that the
European and North American populations of B.
burgdorferi sensu stricto have diverged significantly because of
genetic drift. Plasmid genes evolved independently and showed various effects of
adaptive divergence and diversifying selection. At all 4 loci, genetic
variations within the 2 continents contributed to most (>70%) of the total
sequence diversity, which suggests recent common ancestry, migration, or both,
between the European and North American populations.
Endemic and
Shared ospC Alleles
Gene trees showed
more detailed pictures of geographic variations at each locus (Figure
1). Among the 17 major sequence groups of ospC,
2 minor sequence variants of major-group allele B were geographically distinct
and thereby named B1 in North America and B2 in Europe. Three ospC
alleles (A, E, and K) were observed in both continents, 5 (B2, S, L, Q, and V)
exclusively in Europe (not including the outgroup Ri5 and SV1 alleles), and 10
(B1, C, D, F, G, H, I, J, N, and U) exclusively in North America (Table
1). Although the sample sizes of the North America isolates were small, the
same set of ospC alleles has
repeatedly been identified in surveys of natural populations (1416,38).
These isolates are therefore a reasonably complete representation of ospC
diversity in North America. How well our European samples represent the overall ospC
diversity in Europe is less certain because the European isolates were from an
archived collection rather than from systematic surveys of natural populations.
For instance, ospC alleles J, P,
and R have been identified in Europe (26).
Nonetheless, ospC-A appeared to
be the only allele that is highly prevalent on both continents (Table
1). An earlier study showed that ospC-A
and ospC-B alleles existed in both continents, whereas other ospC
alleles were geographically distinct (K, J, F in North America and P, Q, R, S in
Europe) (24).
Our results further suggested that the ospC-B
clonal group had 2 geographically distinct subtypes (Figure
1, panel B).
Recombinant
Genotypes
Previous MLST
studies showed complete linkage between ospC
and other loci on plasmids or the main chromosome in the North American
populations (15,16).
This finding is consistent with our study, in which allelic types at IGS, dbpA,
and BBD14 of the 68 isolates were almost entirely predictable from their ospC
types. Because of the nearly complete linkage between ospC
and a locus, individual clonal complexes could conveniently be named after their
ospC alleles. However, 5 isolates showed alleles at non-ospC
loci inconsistent with allelic types typically associated with their ospC
alleles, including MI409, MI415, and MI418 from the midwestern United States and
Bol26 and VS219 from Europe (Figure
1). Because these genotypes were new combinations of allelic types found
elsewhere, they are more likely to be recombinant genotypes caused by plasmid
exchanges, rather than locally evolved new genotypes (17).
Notably, these probable recombinants were from samples from either the
midwestern United States or Europe, and none were from the intensively surveyed
northeastern United States. A higher number of clones in the northeastern United
States than elsewhere could be understood because B.
burgdorferi populations in that region are evolutionarily young and
show an epidemic population structure (15,19).
On the basis of the presence of allele types at 4 loci, we determined
preliminarily that Bol26 is a group Q or V clone with a transferred ospC-S
allele because Bol26 clustered with group Q and V isolates at IGS, dbpA,
and BBD14 (Figure
1). By the same reasoning, VS219 is a group B2 clone with a transferred
BBD14 allele. We are currently investigating the donor and recipient genomic
types of these recombinant isolates by sequencing 6 additional loci.
Recent
Trans-Oceanic Dispersals
Three clonal
complexes (A, E, K) are distributed in both continents (Table
1). For the A clonal group, 6 isolates from the United States and 11
isolates from Europe were sequenced at 4 loci. The 4-locus sequences of the
isolates between the 2 continents were identical (Figure
1). Thus, the A clonal complex likely was dispersed across the Atlantic
Ocean rather recently. To verify the genetic homogeneity of group A isolates
from the 2 continents, we randomly selected 4 group A isolates (B31 and 132b
from the United States; IP1 and PKa2 from Europe) for further sequencing at an
additional 6 chromosomal loci. No fixed sequence differences between 2
continental samples were found, which lends further support for the recent
trans-oceanic migration of the A clone (Figure
2). Similarly, the 4-loci sequences of E and K isolates between the 2
continental samples were identical, indicating recent migration of these clonal
groups as well (Figure
1). However, the E and K groups seemed less prevalent in Europe than the A
group (Table 1). Because individual ticks and hosts
are commonly infected with multiple B.
burgdorferi clones, any migration, whether by natural or
human-facilitated mechanisms, is likely to involve a mixture of clonal groups,
rather than a single clone. Upon their arrival, however, clonal groups may
differ in their ability to colonize a new niche consisting of novel vector and
host species. By this reasoning, the A clone is the most ecologically successful
strain, able to thrive in a new niche with little genetic change. This
conclusion is supported by surveys that showed a broad range of host species for
this clonal group (9,10).
We could not
determine conclusively the direction, timing, or number of the trans-oceanic
dispersals. Assuming that the chromosomal gene tree in Figure
2, panel B, is an accurate representation of the phylogeny of these clonal
groups, a parsimonious scenario is that an early migrant from Europe was the
ancestor of the North American clade consisting of the A and B1 groups, and a
more recent migration has introduced the A group to Europe. However, none of the
basal branches of this gene tree was well supported (Figure
2). Multilocus sequencing of more loci, especially rapidly evolving plasmid
loci, of group A isolates will help find more conclusive answers to these
questions. To estimate the time of the A clone migration, we noted that no fixed
differences in nucleotides occurred within a total of 11,167 aligned bases at 7
chromosomal and 3 plasmid loci. If one assumes a neutral evolutionary rate on
the order of 1 substitution per site per million years, the Poisson zero-term
probabilities that no fixed difference has occurred within 11,167 bases in the
past 50, 100, and 200 years are 0.33, 0.10, and 0.011, respectively. Therefore,
the trans-oceanic migration of clone A likely occurred more recently than 200
years ago. More realistic estimates would depend on studies of the neutral
mutation rate and generation time of B.
burgdorferi in the wild.
Phylogenetic
Heterogeneity of Group B Isolates
The ospC-B
clonal group is another highly virulent strain identified by association studies
(2022,24).
Initially, group B seemed to be another clone that is distributed in both
continents with a few sequence differences at IGS and ospC
(Figure 1).
Sequencing at additional 6 housekeeping loci, however, showed deep phylogenetic
heterogeneity of the B group, while the A group remained homogeneous (Figure
2). The 2 B clonal complexes (B1 in North America and B2 in Europe) do not
form a monophyletic clade (Figure
2). Rather, B2 clusters with other European clones (V and Q). Also, clones
B1 and A, the 2 closest North American relatives, do not form a well-supported
clade (only 51% bootstrap support). Clearly, unlike the A clone, the
bicontinental distribution of the B clone is not due to recent migration.
Sharing of similar ospC B
alleles between the 2 continents may be due to stabilizing selection or lateral
transfer. Because few synonymous changes have occurred between the B1 and B2
alleles, lateral transfer is a more likely cause.
The B2/Q/V showed
as a European clade with nearly uniform chromosomal sequences, although it had
highly divergent ospC alleles (Figure
2). This evidence, based on chromosome-wide genes, strengthens the
conclusions of an earlier study that adaptive, large sequence variations at ospC
are associated with incipient genome divergence (19).
Finally, the
overall genospecies phylogeny based on MLST showed that the 2 European isolates
(Ri5 and SV1) that we used as outgroups may be a new genospecies (Figure
2). This phylogeny is robust because tests of recombination using GENECONV
showed no statistically significant gene conversion within the 6 chromosomal
housekeeping loci (37).
The hypothetical genospecies represented by Ri5 and SV1 is more closely related
to B. burgdorferi sensu stricto
than B. bissettii (represented
here by DN127) is to B. burgdorferi sensu
stricto. Thus, the MLST phylogeny suggests a possibility that Europe, rather
than North America, may be the origin of B.
burgdorferi sensu stricto, despite a higher contemporary genetic
heterogeneity in North America than in Europe.
Conclusions
To summarize, the
present study used 7 chromosomal loci (IGS and 6 housekeeping genes) to
reconstruct the intra- and interspecific phylogeographic histories of B.
burgdorferi sensu stricto. Although the standard MLST scheme based
on housekeeping genes enables estimates of recombination and mutation rates as
well as intraspecific phylogenies (2,5),
our approach of including plasmidborne loci under positive selection helped
identify the selective causes of bacterial lineage divergence. Our results
showed significant endemic lineage diversification among regional populations,
discovered recombinant genotypes, and strongly indicated migrations between
North American and European populations in modern times. The highly pathogenic
clonal complex A has a prominent presence in both continents, which suggests its
success in finding ecologic niches that enable it to infect a broad range of
host and vector species. The same genetic basis of the ecologic invasiveness of
the ospC-A clone may be
underlying its high virulence to humans. The emergence of Lyme disease in North
America since the 1970s has been attributed to an increasing overlap of human
and B. burgdorferi habitats (39).
On the basis of our evidence of migration events, we propose that the
trans-oceanic dispersal and colonization of ecologically highly successful
clonal complexes (e.g., the A group) may also have played a substantial role.
Acknowledgments
We acknowledge
the Borrelia sequencing team of
Sherwood R. Casjens, John J. Dunn, Benjamin J. Luft, Claire M. Fraser, Weigang
Qiu, and Steven E. Schutzer, working under grants from the Lyme Disease
Association and National Institutes of Health (AI37256 and AI49003), for access
to unpublished sequence information.
Other supports
from this work include grants GM083722-01 (toW.-G.Q.) and RR03037 (to Hunter
College) from the National Institutes of Health.
Dr Qiu is an
assistant professor in the Department of Biological Sciences at Hunter College
and the Biology Department in the Graduate Center of the City University of New
York. His research interests include the evolution and population biology of
infectious diseases, comparative genomics, and bioinformatics tool development.
References
1.Maiden
MC, Bygraves JA, Feil E, Morelli G, Russell JE, Urwin R, et al. Multilocus
sequence typing: a portable approach to the identification of clones within
populations of pathogenic microorganisms. Proc Natl Acad Sci U S A.
1998;95:31405.
PubMedDOI
2.Feil EJ. Small change: keeping pace with microevolution. Nat Rev
Microbiol. 2004;2:48395.
PubMedDOI
3.Cohan FM. Concepts of bacterial biodiversity for the age of
genomics. In: Fraser CM, Read TD, Nelson KE, editors. Microbial genomes. Totwa
(NJ): Humana Press, Inc.; 2004. p. 17594.
4.Fraser C, Hanage WP, Spratt BG. Neutral microepidemic evolution of
bacterial pathogens. Proc Natl Acad Sci U S A. 2005;102:196873.
PubMedDOI
11.Rauter
C, Hartung T. Prevalence of Borrelia
burgdorferi sensu lato genospecies in Ixodes
ricinus ticks in Europe: a metaanalysis. Appl Environ Microbiol.
2005;71:720316.
PubMedDOI
14.Alghaferi MY, Anderson JM, Park J, Auwaerter PG, Aucott JN, Norris
DE, et al. Borrelia burgdorferi ospC
heterogeneity among human and murine isolates from a defined region of northern
Maryland and southern Pennsylvania: lack of correlation with invasive and
noninvasive genotypes. J Clin Microbiol. 2005;43:187984.
PubMedDOI
16.Bunikis J, Garpmo U, Tsao J, Berglund J, Fish D, Barbour AG.
Sequence typing reveals extensive strain diversity of the Lyme borreliosis
agents Borrelia burgdorferi in
North America and Borrelia afzelii
in Europe. Microbiology. 2004;150:174155.
PubMedDOI
17.Qiu WG, Schutzer SE, Bruno JF, Attie O, Xu Y, Dunn JJ, et al.
Genetic exchange and plasmid transfers in Borrelia
burgdorferi sensu stricto revealed by three-way genome comparisons
and multilocus sequence typing. Proc Natl Acad Sci U S A. 2004;101:141505. PubMedDOI
18.Stevenson B, Miller JC. Intra- and interbacterial genetic exchange
of Lyme disease spirochete erp
genes generates sequence identity amidst diversity. J Mol Evol. 2003;57:30924. PubMedDOI
19.Attie O, Bruno JF, Xu Y, Qiu D, Luft BJ, Qiu WG. Co-evolution of
the outer surface protein C gene (ospC)
and intraspecific lineages of Borrelia
burgdorferi sensu stricto in the northeastern United States. Infect
Genet Evol. 2007;7:112. PubMedDOI
20.Wormser GP, Liveris D, Nowakowski J, Nadelman RB, Cavaliere LF,
McKenna D, et al. Association of specific subtypes of Borrelia
burgdorferi with hematogenous dissemination in early Lyme disease. J
Infect Dis. 1999;180:7205.
PubMedDOI
21.Jones
KL, Glickstein LJ, Damle N, Sikand VK, McHugh G, Steere AC. Borrelia
burgdorferi genetic markers and disseminated disease in patients
with early Lyme disease. J Clin Microbiol. 2006;44:440713.
PubMedDOI
23.Baranton G, Seinost G, Theodore G, Postic D, Dykhuizen D. Distinct
levels of genetic diversity of Borrelia
burgdorferi are associated with different aspects of pathogenicity.
Res Microbiol. 2001;152:14956.
PubMedDOI
24.Lagal V, Postic D, Ruzic-Sabljic E, Baranton G. Genetic diversity
among Borrelia strains
determined by single-strand conformation polymorphism analysis of the ospC
gene and its association with invasiveness. J Clin Microbiol. 2003;41:505965.
PubMedDOI
31.Thompson
JD, Higgins DG, Gibson TJ. CLUSTAL W: improving the sensitivity of progressive
multiple sequence alignment through sequence weighting, position-specific gap
penalties and weight matrix choice. Nucleic Acids Res. 1994;22:467380.
PubMedDOI
32.Huelsenbeck JP, Ronquist F. MRBAYES: Bayesian inference of
phylogenetic trees. Bioinformatics. 2001;17:7545.
PubMedDOI
33.Felsenstein J. PHYLIPphylogeny inference package. Cladistics.
1989;5:1646.
34.Paradis E, Claude J, Strimmer K. APE: analyses of phylogenetics and
evolution in R language. Bioinformatics. 2004;20:28990.
PubMedDOI
35.Excoffier L, Laval G, Schneider S. Arlequin ver 3.0: an integratred
software package for population data analysis. Evol Bioinform Online.
2005;1:4750.
36.Glockner G, Lehmann R, Romualdi A, Pradella S, Schulte-Spechtel U,
Schilhabel M, et al. Comparative analysis of the Borrelia garinii genome. Nucleic Acids Res. 2004;32:603846.
PubMedDOI
38.Wang G, van Dam AP, Dankert J. Evidence for frequent OspC
gene transfer between Borrelia valaisiana
sp. nov. and other Lyme disease spirochetes. FEMS Microbiol Lett.
1999;177:28996.
PubMedDOI
39.Barbour AG, Fish D. The biological and social phenomenon of Lyme
disease. Science. 1993;260:16106.
PubMedDOI
Figures
Figure
1. Gene trees showing nucleotide sequence clusters of 68 Borrelia burgdorferi isolates at 1 chromosomal locus... Figure 2. Species
phylogeny based on concatenated sequences at housekeeping loci...
Tables
Table
1. Borrelia burgdorferi
isolates
Wide Distribution of a High-Virulence Borrelia burgdorferi Clone in
Europe and North America
Wei-Gang Qiu,* John F. Bruno,† William D. McCaig,* Yun Xu,† Ian
Livey,‡ Martin E. Schriefer,§ and Benjamin J. Luft†
*Hunter College of the City University of New York, New York, New York, USA;
†Stony Brook University, Stony Brook, New York, USA; ‡Baxter Innovations
GmBH, Orth/Donau, Austria; and §Centers for Disease Control and Prevention,
Fort Collins, Colorado, USA
Vol. 14, No. 7 • July 2008
*ospC, outer surface
protein C; US, United States; EU, European Union.
†Isolates subjected to
multilocus sequence typing analysis.
‡Type names follow (13),
except that B was split to B1 and B2, and 3 new types (V, X, W) were
assigned to European isolates.
§Number and geographic origins
of an ospC type in our collection.
¶Isolates showing evidence for
plasmid-chromosome recombination.
Table 2. Genomic
markers and PCR primers
Wide Distribution of a High-Virulence Borrelia burgdorferi Clone in
Europe and North America
Wei-Gang Qiu,* John F. Bruno,† William D. McCaig,* Yun Xu,† Ian
Livey,‡ Martin E. Schriefer,§ and Benjamin J. Luft†
*Hunter College of the City University of New York, New York, New York, USA;
†Stony Brook University, Stony Brook, New York, USA; ‡Baxter Innovations
GmBH, Orth/Donau, Austria; and §Centers for Disease Control and Prevention,
Fort Collins, Colorado, USA
Vol. 14, No. 7 • July 2008
Wide Distribution of a High-Virulence Borrelia burgdorferi Clone in
Europe and North America
Wei-Gang Qiu,* John F. Bruno,† William D. McCaig,* Yun Xu,† Ian
Livey,‡ Martin E. Schriefer,§ and Benjamin J. Luft†
*Hunter College of the City University of New York, New York, New York, USA;
†Stony Brook University, Stony Brook, New York, USA; ‡Baxter Innovations
GmBH, Orth/Donau, Austria; and §Centers for Disease Control and Prevention,
Fort Collins, Colorado, USA
Vol. 14, No. 7 • July 2008
Table 3.
Analysis of molecular variance results*†
Locus
Molecular
variance, %
Nucleotide
diversity, π
Fixation index
(FST)‡
Between continents
Within continents
North America
Europe
IGS
19.5
80.5
0.0253
0.0243
0.1952§
ospC
3.13
96.87
0.2066
0.1900
0.0313¶
dbpA
26.5
73.5
0.1480
0.0999
0.2650§
BBD14
2.54
97.46
0.0834
0.1333
0.0254 (NS)
*IGS, intergenic
spacer; ospC, outer surface protein C; NS, not significant
(p>0.05).
†Results were obtained by
using Arlequin 3.1 (35). Samples were 66 IGS
sequences divided into 2 continental populations: North America (36
sequences from New York, Connecticut, Massachusetts, and Michigan) and
Europe (30 sequences from Italy, Austria, France, Germany, Switzerland,
Poland, Hungary, Slovenia, and Finland). Two outgroup sequences (SV1 and
Ri5) were excluded from the European sample. Genetic distances between
haplotypes were based on the Kimura 2-parameter model.
‡Levels of significance were
obtained by 1,000 permutations.
§p<0.001.
¶0.01<p<0.05.
References
Maiden MC, Bygraves JA, Feil E, Morelli G, Russell JE,
Urwin R, et al. Multilocus sequence typing: a portable approach to the
identification of clones within populations of pathogenic microorganisms.
Proc Natl Acad Sci U S A. 1998;95:3140–5.
PubMedDOI
Feil EJ. Small change: keeping pace with microevolution. Nat Rev Microbiol.
2004;2:483–95.
PubMedDOI
Cohan FM. Concepts of bacterial biodiversity for the age of genomics. In:
Fraser CM, Read TD, Nelson KE, editors. Microbial genomes. Totwa (NJ):
Humana Press, Inc.; 2004. p. 175–94.
Fraser C, Hanage WP, Spratt BG. Neutral microepidemic evolution of
bacterial pathogens. Proc Natl Acad Sci U S A. 2005;102:1968–73.
PubMedDOI
Rauter C, Hartung T. Prevalence of Borrelia
burgdorferi sensu lato genospecies in Ixodes ricinus ticks in
Europe: a metaanalysis. Appl Environ Microbiol. 2005;71:7203–16.
PubMedDOI
Alghaferi MY, Anderson JM, Park J, Auwaerter PG, Aucott JN, Norris DE, et
al. Borrelia burgdorferi ospC heterogeneity among human and murine
isolates from a defined region of northern Maryland and southern
Pennsylvania: lack of correlation with invasive and noninvasive genotypes. J
Clin Microbiol. 2005;43:1879–84.
PubMedDOI
Bunikis J, Garpmo U, Tsao J, Berglund J, Fish D, Barbour AG. Sequence
typing reveals extensive strain diversity of the Lyme borreliosis agents Borrelia
burgdorferi in North America and Borrelia afzelii in Europe.
Microbiology. 2004;150:1741–55.
PubMedDOI
Qiu WG, Schutzer SE, Bruno JF, Attie O, Xu Y, Dunn JJ, et al. Genetic
exchange and plasmid transfers in Borrelia burgdorferi sensu
stricto revealed by three-way genome comparisons and multilocus sequence
typing. Proc Natl Acad Sci U S A. 2004;101:14150–5. PubMedDOI
Stevenson B, Miller JC. Intra- and interbacterial genetic exchange of Lyme
disease spirochete erp genes generates sequence identity amidst
diversity. J Mol Evol. 2003;57:309–24. PubMedDOI
Attie O, Bruno JF, Xu Y, Qiu D, Luft BJ, Qiu WG. Co-evolution of the outer
surface protein C gene (ospC) and intraspecific lineages of Borrelia
burgdorferi sensu stricto in the northeastern United States. Infect
Genet Evol. 2007;7:1–12. PubMedDOI
Wormser GP, Liveris D, Nowakowski J, Nadelman RB, Cavaliere LF, McKenna D,
et al. Association of specific subtypes of Borrelia burgdorferi
with hematogenous dissemination in early Lyme disease. J Infect Dis.
1999;180:720–5.
PubMedDOI
Jones KL, Glickstein LJ, Damle N, Sikand VK, McHugh G,
Steere AC. Borrelia burgdorferi genetic markers and disseminated
disease in patients with early Lyme disease. J Clin Microbiol.
2006;44:4407–13.
PubMedDOI
Baranton G, Seinost G, Theodore G, Postic D, Dykhuizen D. Distinct levels
of genetic diversity of Borrelia burgdorferi are associated with
different aspects of pathogenicity. Res Microbiol. 2001;152:149–56.
PubMedDOI
Lagal V, Postic D, Ruzic-Sabljic E, Baranton G. Genetic diversity among Borrelia
strains determined by single-strand conformation polymorphism analysis of
the ospC gene and its association with invasiveness. J Clin
Microbiol. 2003;41:5059–65.
PubMedDOI
Thompson JD, Higgins DG, Gibson TJ. CLUSTAL W: improving
the sensitivity of progressive multiple sequence alignment through sequence
weighting, position-specific gap penalties and weight matrix choice. Nucleic
Acids Res. 1994;22:4673–80.
PubMedDOI
Huelsenbeck JP, Ronquist F. MRBAYES: Bayesian inference of phylogenetic
trees. Bioinformatics. 2001;17:754–5.
PubMedDOI
Felsenstein J. PHYLIP—phylogeny inference package. Cladistics.
1989;5:164–6.
Paradis E, Claude J, Strimmer K. APE: analyses of phylogenetics and
evolution in R language. Bioinformatics. 2004;20:289–90.
PubMedDOI
Excoffier L, Laval G, Schneider S. Arlequin ver 3.0: an integratred
software package for population data analysis. Evol Bioinform Online.
2005;1:47–50.
Glockner G, Lehmann R, Romualdi A, Pradella S, Schulte-Spechtel U,
Schilhabel M, et al. Comparative analysis of the Borrelia garinii
genome. Nucleic Acids Res. 2004;32:6038–46.
PubMedDOI
Wang G, van Dam AP, Dankert J. Evidence for frequent OspC gene
transfer between Borrelia valaisiana sp. nov. and other Lyme
disease spirochetes. FEMS Microbiol Lett. 1999;177:289–96.
PubMedDOI
Barbour AG, Fish D. The biological and social phenomenon of Lyme disease.
Science. 1993;260:1610–6.
PubMedDOI
This page posted June 30, 2008
This page last reviewed June 30, 2008
Suggested Citation for this Article
Qiu W-G, Bruno JF,
McCaig WD, Xu Y, Livey I, Schriefer ME, et al. Wide distribution of a
high-virulence Borrelia burgdorferi
clone in Europe and North America. Emerg Infect Dis [serial on the Internet].
2008 Jul [date cited]. Available
from
DOI: 10.3201/eid1407.070880
Comments
to the Authors
Please
contact them at the following address:
Wei-Gang Qiu, Department of Biological Sciences, Hunter College of
the City University of New York, 695 Park Ave, New York, NY 10065, USA; email: weigang@genectr.hunter.cuny.edu
General
Information
USAGov
-- The U.S. Government's Official Web Portal
can be found athttp://www.usa.gov/
FOIA
for the CDC can be found at http://www.cdc.gov/od/foia/index.htm
Department
of Health and Human Services are at http://www.hhs.gov/
Safer Healthier People
This article appeared in the EID Journal Home > Volume
14, Number 7 July 2008 on the Centers
for Disease Control website
Centers
for Disease Control and Prevention, 1600 Clifton Rd, Atlanta, GA 30333, U.S.A
Tel: (404) 639-3311 / Public Inquiries: (404) 639-3534 / (800) 311-3435
EID Editors may be emailed to eideditor@cdc.gov
CDC
Disclaimer:
The opinions expressed by authors contributing to this journal do not
necessarily reflect the opinions of the U.S. Department of Health and Human
Services, the Public Health Service, the Centers for Disease Control and
Prevention, or the authors' affiliated institutions. Use of trade names is for
identification only and does not imply endorsement by any of the groups named
above.www.cdc.gov/ncidod/eid